Black Raspberries and Protocatechuic Acid Mitigate DNFB-Induced Contact Hypersensitivity by Down-Regulating Dendritic Cell Activation and Inhibiting Mediators of Effector Responses

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Handling
2.2. Animal Diets
2.3. Black Raspberry Extract Preparation for In Vitro Studies
2.4. Induction of Contact Hypersensitivity
2.5. Flow Cytometry
2.6. Histopathology
2.7. LPS Stimulation of RAW264.7 Macrophages, and Bone Marrow-Derived Macrophages and Dendritic Cells
2.8. Real Time Quantitative PCR
2.9. Enzyme-Linked Immunosorbent Assay
2.10. Griess and Cytotoxicity Assay
2.11. Statistical Analysis
3. Results and Discussion
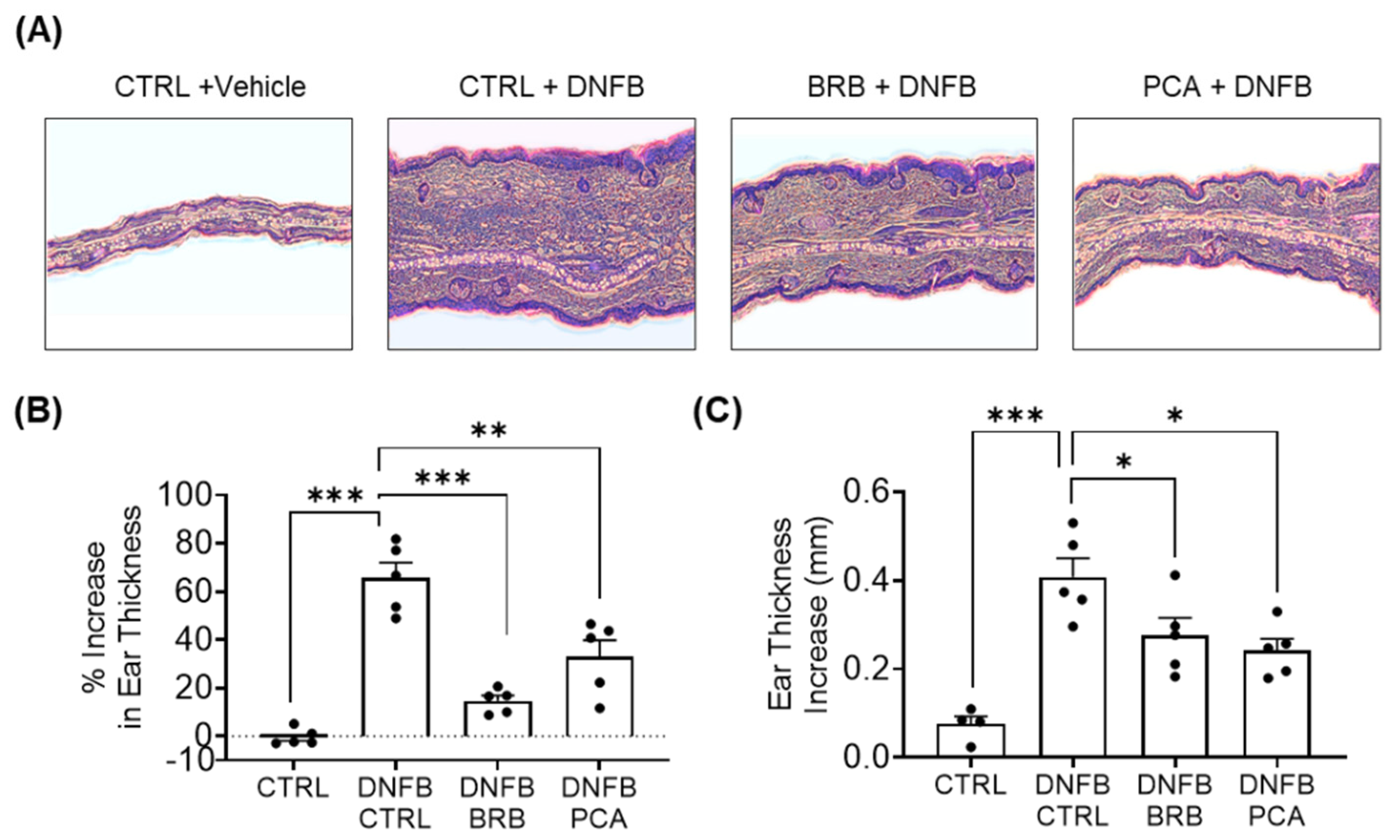
3.1. Auricular Inflammation Is Attenuated in Mice Fed with BRB and PCA Supplemented Diets during DNFB-Induced CHS
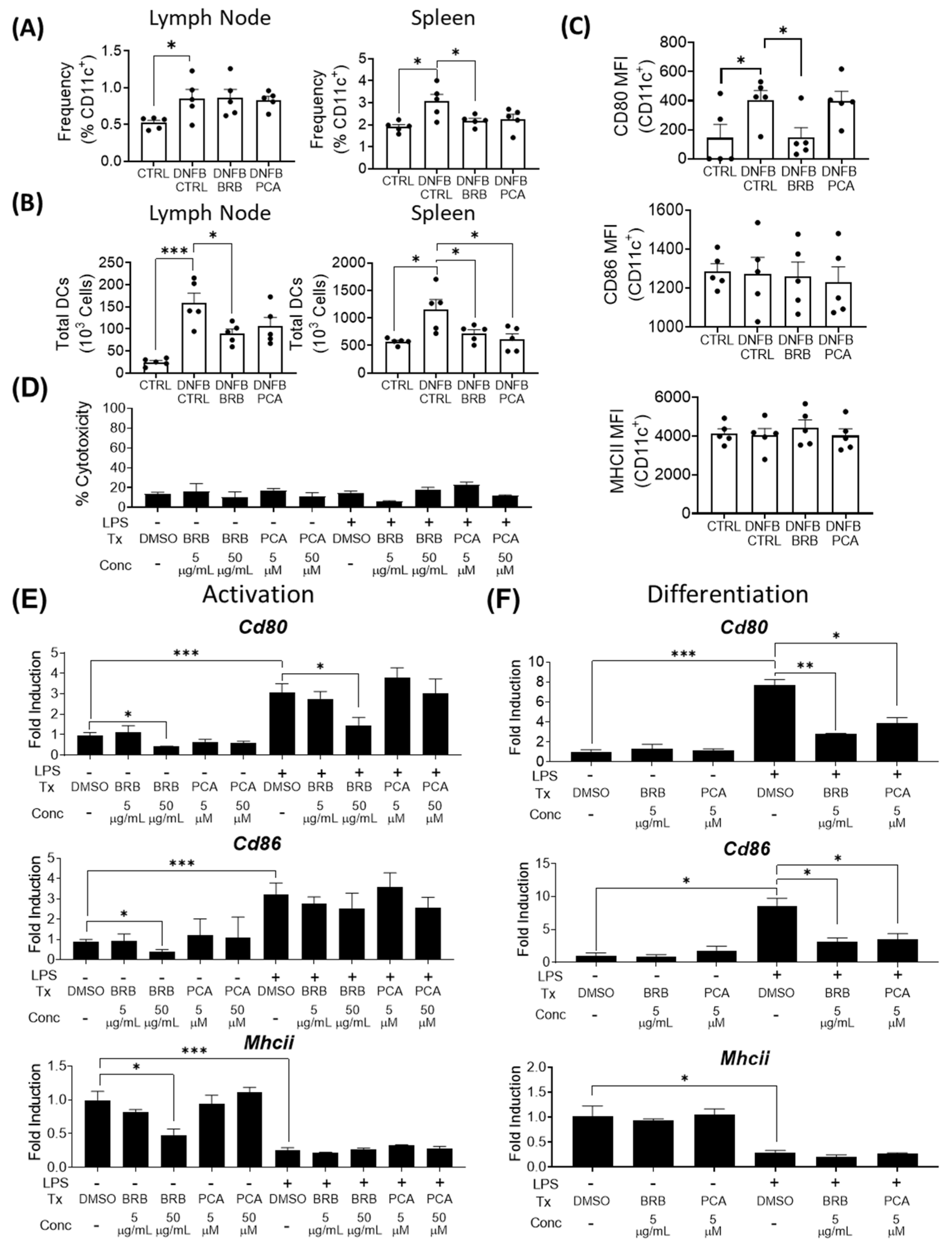
3.2. Effects of BRB and PCA on Dendritic Cell Migration, Maturation and Antigen Presentation during DNFB Induced CHS
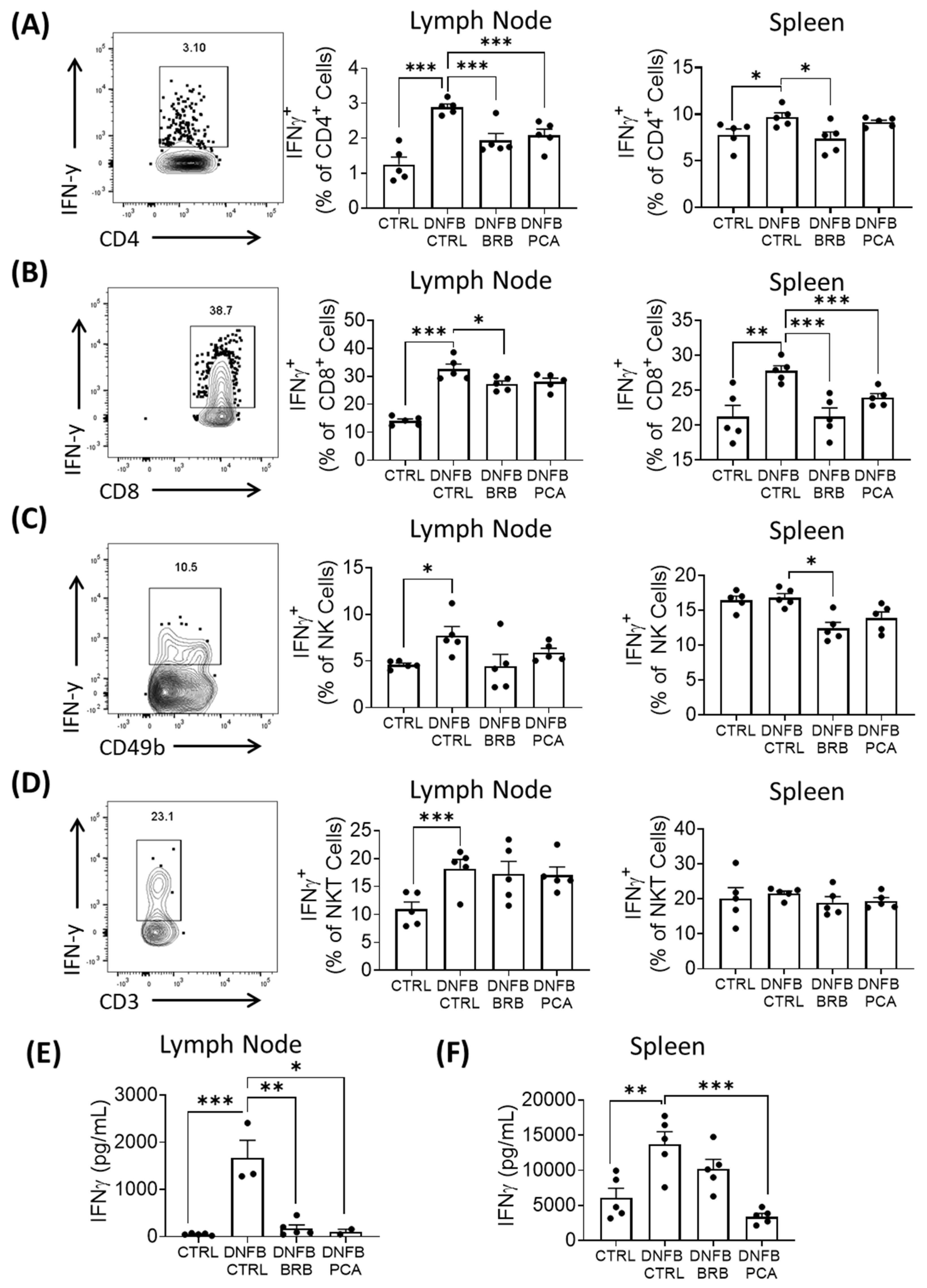
3.3. IL-12, a Key Mediator of DNFB-Induced CHS Is Reduced by BRB-E and PCA
3.4. IFN-γ Production Is Attenuated in DNFB Challenged Mice Fed BRB and, to a Lesser Extent, PCA Supplemented Diets
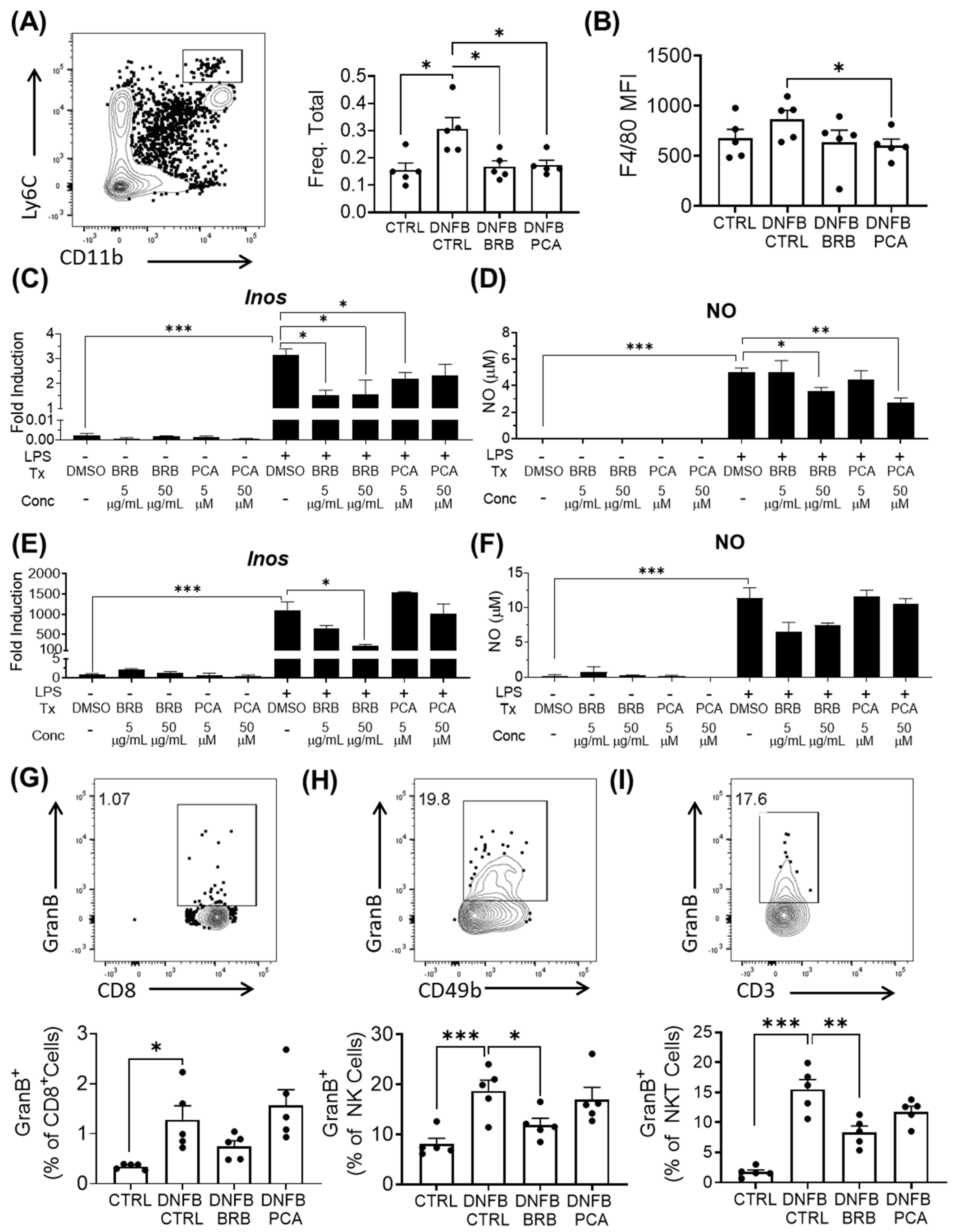
3.5. Mediators of Effector Responses during CHS, Are Inhibited by BRB and PCA
3.6. Il-17 Production Is Significantly Reduced by PCA Supplemented Diet, but Not BRB Supplemented Diet
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Murphy, P.B.; Hooten, J.N.; Atwater, A.R.; Gossman, W. Allergic Contact Dermatitis; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Brasch, J.; Becker, D.; Aberer, W.; Bircher, A.; Kranke, B.; Jung, K.; Przybilla, B.; Biedermann, T.; Werfel, T.; John, S.M.; et al. Guideline contact dermatitis: S1-Guidelines of the German Contact Allergy Group (DKG) of the German Dermatology Society (DDG), the Information Network of Dermatological Clinics (IVDK), the German Society for Allergology and Clinical Immunology (DGAKI), the Working Group for Occupational and Environmental Dermatology (ABD) of the DDG, the Medical Association of German Allergologists (AeDA), the Professional Association of German Dermatologists (BVDD) and the DDG. Allergo J. Int. 2014, 23, 126–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalgard, F.J.; Svensson, A.; Halvorsen, J.A.; Gieler, U.; Schut, C.; Tomas-Aragones, L.; Lien, L.; Poot, F.; Jemec, G.B.E.; Misery, L.; et al. Itch and Mental Health in Dermatological Patients across Europe: A Cross-Sectional Study in 13 Countries. J. Investig. Dermatol. 2020, 140, 568–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peiser, M.; Tralau, T.; Heidler, J.; Api, A.M.; Arts, J.H.; Basketter, D.A.; English, J.; Diepgen, T.L.; Fuhlbrigge, R.C.; Gaspari, A.A.; et al. Allergic contact dermatitis: Epidemiology, molecular mechanisms, in vitro methods and regulatory aspects. Current knowledge assembled at an international workshop at BfR, Germany. Cell Mol. Life Sci. 2012, 69, 763–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usatine, R.P.; Riojas, M. Diagnosis and management of contact dermatitis. Am. Fam. Physician 2010, 82, 249–255. [Google Scholar] [PubMed]
- Yasir, M.; Sonthalia, S. Corticosteroid Adverse Effects; StatPearls: Treasure Island, FL, USA, 2019. [Google Scholar]
- Vocanson, M.; Hennino, A.; Rozieres, A.; Poyet, G.; Nicolas, J.F. Effector and regulatory mechanisms in allergic contact dermatitis. Allergy 2009, 64, 1699–1714. [Google Scholar] [CrossRef] [PubMed]
- Nagamachi, M.; Sakata, D.; Kabashima, K.; Furuyashiki, T.; Murata, T.; Segi-Nishida, E.; Soontrapa, K.; Matsuoka, T.; Miyachi, Y.; Narumiya, S. Facilitation of Th1-mediated immune response by prostaglandin E receptor EP1. J. Exp. Med. 2007, 204, 2865–2874. [Google Scholar] [CrossRef]
- Ogawa, A.; Yoshizaki, A.; Yanaba, K.; Ogawa, F.; Hara, T.; Muroi, E.; Takenaka, M.; Shimizu, K.; Hasegawa, M.; Fujimoto, M.; et al. The differential role of L-selectin and ICAM-1 in Th1-type and Th2-type contact hypersensitivity. J. Investig. Dermatol. 2010, 130, 1558–1570. [Google Scholar] [CrossRef] [Green Version]
- Erkes, D.A.; Selvan, S.R. Hapten-induced contact hypersensitivity, autoimmune reactions, and tumor regression: Plausibility of mediating antitumor immunity. J. Immunol. Res. 2014, 2014, 175265. [Google Scholar] [CrossRef]
- Akiba, H.; Kehren, J.; Ducluzeau, M.T.; Krasteva, M.; Horand, F.; Kaiserlian, D.; Kaneko, F.; Nicolas, J.F. Skin inflammation during contact hypersensitivity is mediated by early recruitment of CD8+ T cytotoxic 1 cells inducing keratinocyte apoptosis. J. Immunol. 2002, 168, 3079–3087. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.H.; Kwon, S.O.; Lee, S.Y.; Kim, H.Y.; Kwon, J.W.; Kim, B.J.; Yu, J.; Kim, H.B.; Kim, W.K.; Jang, G.C.; et al. Association of antioxidants with allergic rhinitis in children from seoul. Allergy Asthma Immunol. Res. 2013, 5, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Gref, A.; Rautiainen, S.; Gruzieva, O.; Hakansson, N.; Kull, I.; Pershagen, G.; Wickman, M.; Wolk, A.; Melen, E.; Bergstrom, A. Dietary total antioxidant capacity in early school age and subsequent allergic disease. Clin. Exp. Allergy 2017, 47, 751–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagano, T.; Katase, M.; Tsumura, K. Impact of soymilk consumption on 2,4-dinitrofluorobenzene-induced contact hypersensitivity and gut microbiota in mice. Int. J. Food Sci. Nutr. 2019, 70, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Nagano, T.; Katase, M.; Tsumura, K. Inhibitory effects of dietary soy isoflavone and gut microbiota on contact hypersensitivity in mice. Food Chem. 2019, 272, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Sakai, S.; Sugawara, T.; Hirata, T. Inhibitory effect of dietary carotenoids on dinitrofluorobenzene-induced contact hypersensitivity in mice. Biosci. Biotechnol. Biochem. 2011, 75, 1013–1015. [Google Scholar] [CrossRef]
- Son, H.U.; Lee, S.; Heo, J.C.; Lee, S.H. The solid-state fermentation of Artemisia capillaris leaves with Ganoderma lucidum enhances the anti-inflammatory effects in a model of atopic dermatitis. Int. J. Mol. Med. 2017, 39, 1233–1241. [Google Scholar] [CrossRef] [Green Version]
- Knobloch, T.J.; Uhrig, L.K.; Pearl, D.K.; Casto, B.C.; Warner, B.M.; Clinton, S.K.; Sardo-Molmenti, C.L.; Ferguson, J.M.; Daly, B.T.; Riedl, K.; et al. Suppression of Proinflammatory and Prosurvival Biomarkers in Oral Cancer Patients Consuming a Black Raspberry Phytochemical-Rich Troche. Cancer Prev. Res. 2016, 9, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Knobloch, T.J.; Ryan, N.M.; Bruschweiler-Li, L.; Wang, C.; Bernier, M.C.; Somogyi, A.; Yan, P.S.; Cooperstone, J.L.; Mo, X.; Brüschweiler, R.P.; et al. Metabolic Regulation of Glycolysis and AMP Activated Protein Kinase Pathways during Black Raspberry-Mediated Oral Cancer Chemoprevention. Metabolites 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Oghumu, S.; Casto, B.C.; Ahn-Jarvis, J.; Weghorst, L.C.; Maloney, J.; Geuy, P.; Horvath, K.Z.; Bollinger, C.E.; Warner, B.M.; Summersgill, K.F.; et al. Inhibition of Pro-inflammatory and Anti-apoptotic Biomarkers during Experimental Oral Cancer Chemoprevention by Dietary Black Raspberries. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Paudel, L.; Wyzgoski, F.J.; Scheerens, J.C.; Chanon, A.M.; Reese, R.N.; Smiljanic, D.; Wesdemiotis, C.; Blakeslee, J.J.; Riedl, K.M.; Rinaldi, P.L. Nonanthocyanin secondary metabolites of black raspberry (Rubus occidentalis L.) fruits: Identification by HPLC-DAD, NMR, HPLC-ESI-MS, and ESI-MS/MS analyses. J. Agric. Food Chem. 2013, 61, 12032–12043. [Google Scholar] [CrossRef]
- Bi, X.; Fang, W.; Wang, L.S.; Stoner, G.D.; Yang, W. Black raspberries inhibit intestinal tumorigenesis in apc1638+/- and Muc2-/- mouse models of colorectal cancer. Cancer Prev. Res. 2010, 3, 1443–1450. [Google Scholar] [CrossRef] [Green Version]
- Harris, G.K.; Gupta, A.; Nines, R.G.; Kresty, L.A.; Habib, S.G.; Frankel, W.L.; LaPerle, K.; Gallaher, D.D.; Schwartz, S.J.; Stoner, G.D. Effects of lyophilized black raspberries on azoxymethane-induced colon cancer and 8-hydroxy-2′-deoxyguanosine levels in the Fischer 344 rat. Nutr. Cancer 2001, 40, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Stoner, G.D.; Wang, L.S.; Casto, B.C. Laboratory and clinical studies of cancer chemoprevention by antioxidants in berries. Carcinogenesis 2008, 29, 1665–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.S.; Hecht, S.S.; Carmella, S.G.; Yu, N.; Larue, B.; Henry, C.; McIntyre, C.; Rocha, C.; Lechner, J.F.; Stoner, G.D. Anthocyanins in black raspberries prevent esophageal tumors in rats. Cancer Prev. Res. 2009, 2, 84–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keppler, K.; Humpf, H.U. Metabolism of anthocyanins and their phenolic degradation products by the intestinal microflora. Bioorg. Med. Chem. 2005, 13, 5195–5205. [Google Scholar] [CrossRef] [PubMed]
- Guttenplan, J.B.; Chen, K.M.; Sun, Y.W.; Kosinska, W.; Zhou, Y.; Kim, S.A.; Sung, Y.; Gowda, K.; Amin, S.; Stoner, G.D.; et al. Effects of Black Raspberry Extract and Protocatechuic Acid on Carcinogen-DNA Adducts and Mutagenesis, and Oxidative Stress in Rat and Human Oral Cells. Cancer Prev. Res. 2016, 9, 704–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peiffer, D.S.; Zimmerman, N.P.; Wang, L.S.; Ransom, B.W.; Carmella, S.G.; Kuo, C.T.; Siddiqui, J.; Chen, J.H.; Oshima, K.; Huang, Y.W.; et al. Chemoprevention of esophageal cancer with black raspberries, their component anthocyanins, and a major anthocyanin metabolite, protocatechuic acid. Cancer Prev. Res. 2014, 7, 574–584. [Google Scholar] [CrossRef] [Green Version]
- Stoner, G.D. Foodstuffs for preventing cancer: The preclinical and clinical development of berries. Cancer Prev. Res. 2009, 2, 187–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.; Thomas-Ahner, J.M.; Riedl, K.M.; Bailey, M.T.; Vodovotz, Y.; Schwartz, S.J.; Clinton, S.K. Dietary Black Raspberries Impact the Colonic Microbiome and Phytochemical Metabolites in Mice. Mol. Nutr. Food Res. 2019, 63, e1800636. [Google Scholar] [CrossRef]
- Wei, M.; Chu, X.; Guan, M.; Yang, X.; Xie, X.; Liu, F.; Chen, C.; Deng, X. Protocatechuic acid suppresses ovalbumin-induced airway inflammation in a mouse allergic asthma model. Int. Immunopharmacol. 2013, 15, 780–788. [Google Scholar] [CrossRef]
- Wu, X.; Pittman Iii, H.E.; Hager, T.; Hager, A.; Howard, L.; Prior, R.L. Phenolic acids in black raspberry and in the gastrointestinal tract of pigs following ingestion of black raspberry. Mol. Nutr. Food Res. 2009, 53 (Suppl. 1), S76–S84. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and Measurement of Anthocyanins by UV-Visible Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, F1.2.1–F1.2.13. [Google Scholar] [CrossRef]
- Report of the American Institute of Nurtition ad hoc Committee on Standards for Nutritional Studies. J. Nutr. 1977, 107, 1340–1348. [CrossRef]
- Gaspari, A.A.; Katz, S.I.; Martin, S.F. Contact Hypersensitivity. Curr. Protoc. Immunol. 2016, 113, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, G.; Terrazas, C.; Oghumu, S.; Varikuti, S.; Dubovsky, J.A.; Byrd, J.C.; Satoskar, A.R. Ibrutinib enhances IL-17 response by modulating the function of bone marrow derived dendritic cells. Oncoimmunology 2016, 5, e1057385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, Y.; Aiba, S. Dendritic cells and contact dermatitis. Clin. Rev. Allergy Immunol. 2007, 33, 27–34. [Google Scholar] [CrossRef]
- Jackson, D.G. Leucocyte Trafficking via the Lymphatic Vasculature-Mechanisms and Consequences. Front. Immunol. 2019, 10, 471. [Google Scholar] [CrossRef]
- Pulgaonkar, R.; Chitra, P.; Moosvi, Z. Benign reactive lymphadenopathy associated with submandibular gland enlargement during orthodontic treatment. J. Oral Maxillofac. Pathol. 2017, 21, 181. [Google Scholar] [CrossRef]
- Katayama, I.; Matsunaga, T.; Yokozeki, H.; Nishioka, K. Blockade of costimulatory molecules B7-1 (CD80) and B7-2 (CD86) down-regulates induction of contact sensitivity by haptenated epidermal cells. Br. J. Derm. 1997, 136, 846–852. [Google Scholar] [CrossRef]
- Yu, X.; Fukunaga, A.; Nagai, H.; Oniki, S.; Honma, N.; Ichihashi, M.; Matozaki, T.; Nishigori, C.; Horikawa, T. Engagement of CD47 inhibits the contact hypersensitivity response via the suppression of motility and B7 expression by Langerhans cells. J. Investig. Dermatol. 2006, 126, 797–807. [Google Scholar] [CrossRef] [Green Version]
- Ushio, H.; Tsuji, R.F.; Szczepanik, M.; Kawamoto, K.; Matsuda, H.; Askenase, P.W. IL-12 reverses established antigen-specific tolerance of contact sensitivity by affecting costimulatory molecules B7-1 (CD80) and B7-2 (CD86). J. Immunol. 1998, 160, 2080–2088. [Google Scholar]
- Henry, C.J.; Ornelles, D.A.; Mitchell, L.M.; Brzoza-Lewis, K.L.; Hiltbold, E.M. IL-12 produced by dendritic cells augments CD8+ T cell activation through the production of the chemokines CCL1 and CCL17. J. Immunol. 2008, 181, 8576–8584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Mizoguchi, I.; Morishima, N.; Chiba, Y.; Mizuguchi, J.; Yoshimoto, T. Regulation of antitumor immune responses by the IL-12 family cytokines, IL-12, IL-23, and IL-27. Clin. Dev. Immunol. 2010, 2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riemann, H.; Schwarz, A.; Grabbe, S.; Aragane, Y.; Luger, T.A.; Wysocka, M.; Kubin, M.; Trinchieri, G.; Schwarz, T. Neutralization of IL-12 in vivo prevents induction of contact hypersensitivity and induces hapten-specific tolerance. J. Immunol. 1996, 156, 1799–1803. [Google Scholar]
- Trinchieri, G.; Gerosa, F. Immunoregulation by interleukin-12. J. Leukoc. Biol. 1996, 59, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Yan, W.; Zheng, H.; Du, Q.; Zhang, L.; Ban, Y.; Li, N.; Wei, F. Regulation of IL-10 and IL-12 production and function in macrophages and dendritic cells. F1000Research 2015, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Athie-Morales, V.; Smits, H.H.; Cantrell, D.A.; Hilkens, C.M. Sustained IL-12 signaling is required for Th1 development. J. Immunol. 2004, 172, 61–69. [Google Scholar] [CrossRef]
- Heufler, C.; Koch, F.; Stanzl, U.; Topar, G.; Wysocka, M.; Trinchieri, G.; Enk, A.; Steinman, R.M.; Romani, N.; Schuler, G. Int.erleukin-12 is produced by dendritic cells and mediates T helper 1 development as well as interferon-gamma production by T helper 1 cells. Eur. J. Immunol. 1996, 26, 659–668. [Google Scholar] [CrossRef]
- He, D.; Wu, L.; Kim, H.K.; Li, H.; Elmets, C.A.; Xu, H. IL-17 and IFN-gamma mediate the elicitation of contact hypersensitivity responses by different mechanisms and both are required for optimal responses. J. Immunol. 2009, 183, 1463–1470. [Google Scholar] [CrossRef] [Green Version]
- Paust, S.; Senman, B.; von Andrian, U.H. Adaptive immune responses mediated by natural killer cells. Immunol. Rev. 2010, 235, 286–296. [Google Scholar] [CrossRef]
- Miller, G.; Lahrs, S.; Dematteo, R.P. Overexpression of interleukin-12 enables dendritic cells to activate NK cells and confer systemic antitumor immunity. FASEB J. 2003, 17, 728–730. [Google Scholar] [CrossRef]
- Luque, I.; Reyburn, H.; Strominger, J.L. Expression of the CD80 and CD86 molecules enhances cytotoxicity by human natural killer cells. Hum. Immunol. 2000, 61, 721–728. [Google Scholar] [CrossRef]
- Guckel, B.; Meyer, G.C.; Rudy, W.; Batrla, R.; Meuer, S.C.; Bastert, G.; Wallwiener, D.; Moebius, U. Int.erleukin-12 requires initial CD80-mediated T-cell activation to support immune responses toward human breast and ovarian carcinoma. Cancer Gene Ther. 1999, 6, 228–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rustemeyer, T.; van Hoogstraten, I.M.W.; von Blomberg, B.M.E.; Scheper, R.J. Mechanisms of Allergic Contact Dermatitis. In Kanerva′s Occupational Dermatology; Rustemeyer, T., Elsner, P., John, S.-M., Maibach, H.I., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 113–146. [Google Scholar] [CrossRef]
- Honda, T.; Egawa, G.; Grabbe, S.; Kabashima, K. Update of immune events in the murine contact hypersensitivity model: Toward the understanding of allergic contact dermatitis. J. Investig. Dermatol. 2013, 133, 303–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, S.Z.; Tan, K.W.; Wong, F.H.S.; Chua, Y.L.; Tang, Y.; Ng, L.G.; Angeli, V.; Kemeny, D.M. CD8 T cells regulate allergic contact dermatitis by modulating CCR2-dependent TNF/iNOS-expressing Ly6C+ CD11b+ monocytic cells. J. Investig. Dermatol. 2014, 134, 666–676. [Google Scholar] [CrossRef] [Green Version]
- Yun, C.; Jung, Y.; Chun, W.; Yang, B.; Ryu, J.; Lim, C.; Kim, J.H.; Kim, H.; Cho, S.I. Anti-Inflammatory Effects of Artemisia Leaf Extract in Mice with Contact Dermatitis In Vitro and In Vivo. Mediat. Inflamm. 2016, 2016, 8027537. [Google Scholar] [CrossRef] [Green Version]
- Qu, R.; Chen, X.; Hu, J.; Fu, Y.; Peng, J.; Li, Y.; Chen, J.; Li, P.; Liu, L.; Cao, J.; et al. Ghrelin protects against contact dermatitis and psoriasiform skin inflammation by antagonizing TNF-alpha/NF-kappaB signaling pathways. Sci. Rep. 2019, 9, 1348. [Google Scholar] [CrossRef] [Green Version]
- Kurimoto, I.; Grammer, S.F.; Shimizu, T.; Nakamura, T.; Streilein, J.W. Role of F4/80+ cells during induction of hapten-specific contact hypersensitivity. Immunology 1995, 85, 621–629. [Google Scholar]
- Fyhrquist, N.; Wolff, H.; Lauerma, A.; Alenius, H. CD8+ T cell migration to the skin requires CD4+ help in a murine model of contact hypersensitivity. PLoS ONE 2012, 7, e41038. [Google Scholar] [CrossRef] [Green Version]
- Montrose, D.C.; Horelik, N.A.; Madigan, J.P.; Stoner, G.D.; Wang, L.S.; Bruno, R.S.; Park, H.J.; Giardina, C.; Rosenberg, D.W. Anti-inflammatory effects of freeze-dried black raspberry powder in ulcerative colitis. Carcinogenesis 2011, 32, 343–350. [Google Scholar] [CrossRef]
- Mallery, S.R.; Zwick, J.C.; Pei, P.; Tong, M.; Larsen, P.E.; Shumway, B.S.; Lu, B.; Fields, H.W.; Mumper, R.J.; Stoner, G.D. Topical application of a bioadhesive black raspberry gel modulates gene expression and reduces cyclooxygenase 2 protein in human premalignant oral lesions. Cancer Res. 2008, 68, 4945–4957. [Google Scholar] [CrossRef] [Green Version]
- Yawalkar, N.; Hunger, R.E.; Buri, C.; Schmid, S.; Egli, F.; Brand, C.U.; Mueller, C.; Pichler, W.J.; Braathen, L.R. A comparative study of the expression of cytotoxic proteins in allergic contact dermatitis and psoriasis: Spongiotic skin lesions in allergic contact dermatitis are highly infiltrated by T cells expressing perforin and granzyme B. Am. J. Pathol. 2001, 158, 803–808. [Google Scholar] [CrossRef]
- Peiser, M. Role of Th17 cells in skin inflammation of allergic contact dermatitis. Clin. Dev. Immunol. 2013, 2013, 261037. [Google Scholar] [CrossRef] [PubMed]
- Kelepouri, D.; Mavropoulos, A.; Bogdanos, D.P.; Sakkas, L.I. The Role of Flavonoids in Inhibiting Th17 Responses in Inflammatory Arthritis. J. Immunol. Res. 2018, 2018, 9324357. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Grams Per kg of Diet | ||
|---|---|---|---|
| AIN-76A (Control) | AIN-76A + PCA | AIN-76A + BRB | |
| Casein | 200 | 200 | 200 |
| DL-methionine | 3 | 3 | 3 |
| Sucrose | 500 | 449.5 | 450 |
| Cornstarch | 150 | 150 | 150 |
| Corn oil | 50 | 50 | 50 |
| Cellulose | 50 | 50 | 50 |
| Mineral mix a | 35 | 35 | 35 |
| Vitamin mix b | 10 | 10 | 10 |
| Choline bitartrate | 2 | 2 | 2 |
| Protocatechuic acid c | 0 | 0.5 | 0.004 |
| Freeze-dried black raspberry powder d | 0 | 0 | 50 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anderson, K.; Ryan, N.; Siddiqui, A.; Pero, T.; Volpedo, G.; Cooperstone, J.L.; Oghumu, S. Black Raspberries and Protocatechuic Acid Mitigate DNFB-Induced Contact Hypersensitivity by Down-Regulating Dendritic Cell Activation and Inhibiting Mediators of Effector Responses. Nutrients 2020, 12, 1701. https://doi.org/10.3390/nu12061701
Anderson K, Ryan N, Siddiqui A, Pero T, Volpedo G, Cooperstone JL, Oghumu S. Black Raspberries and Protocatechuic Acid Mitigate DNFB-Induced Contact Hypersensitivity by Down-Regulating Dendritic Cell Activation and Inhibiting Mediators of Effector Responses. Nutrients. 2020; 12(6):1701. https://doi.org/10.3390/nu12061701
Chicago/Turabian StyleAnderson, Kelvin, Nathan Ryan, Arham Siddiqui, Travis Pero, Greta Volpedo, Jessica L. Cooperstone, and Steve Oghumu. 2020. "Black Raspberries and Protocatechuic Acid Mitigate DNFB-Induced Contact Hypersensitivity by Down-Regulating Dendritic Cell Activation and Inhibiting Mediators of Effector Responses" Nutrients 12, no. 6: 1701. https://doi.org/10.3390/nu12061701