
Association of Plant-Based and High-Protein Diets with a Lower Obesity Risk Defined by Fat Mass in Middle-Aged and Elderly Persons with a High Genetic Risk of Obesity

Abstract
:1. Introduction
2. Methods
2.1. Participants and Setting
2.2. Demographic, Anthropometric, and Biochemical Parameters of the Participants
2.3. Definition of Obesity and Metabolic Syndrome
2.4. Usual Food Intake Using a Semi-Quantitative Food Frequency Questionnaire (SQFFQ)
2.5. Dietary Patterns by Principal Components Analysis
2.6. Dietary Inflammatory Index (DII)
2.7. Genotyping Using a Korean CHIP and Quality Control
2.8. Selection of the Genetic Variants That Influence Obesity Defined by Fat Mass and the Best Model with SNP–SNP Interactions
2.9. Expression Quantitative Trait Locus (eQTL) Analysis
2.10. Statistical Analysis
3. Results
3.1. Demographic Characteristics and Lifestyles According to Genders and Obesity
3.2. Anthropometric and Biochemical Parameters According to Genders and Obesity
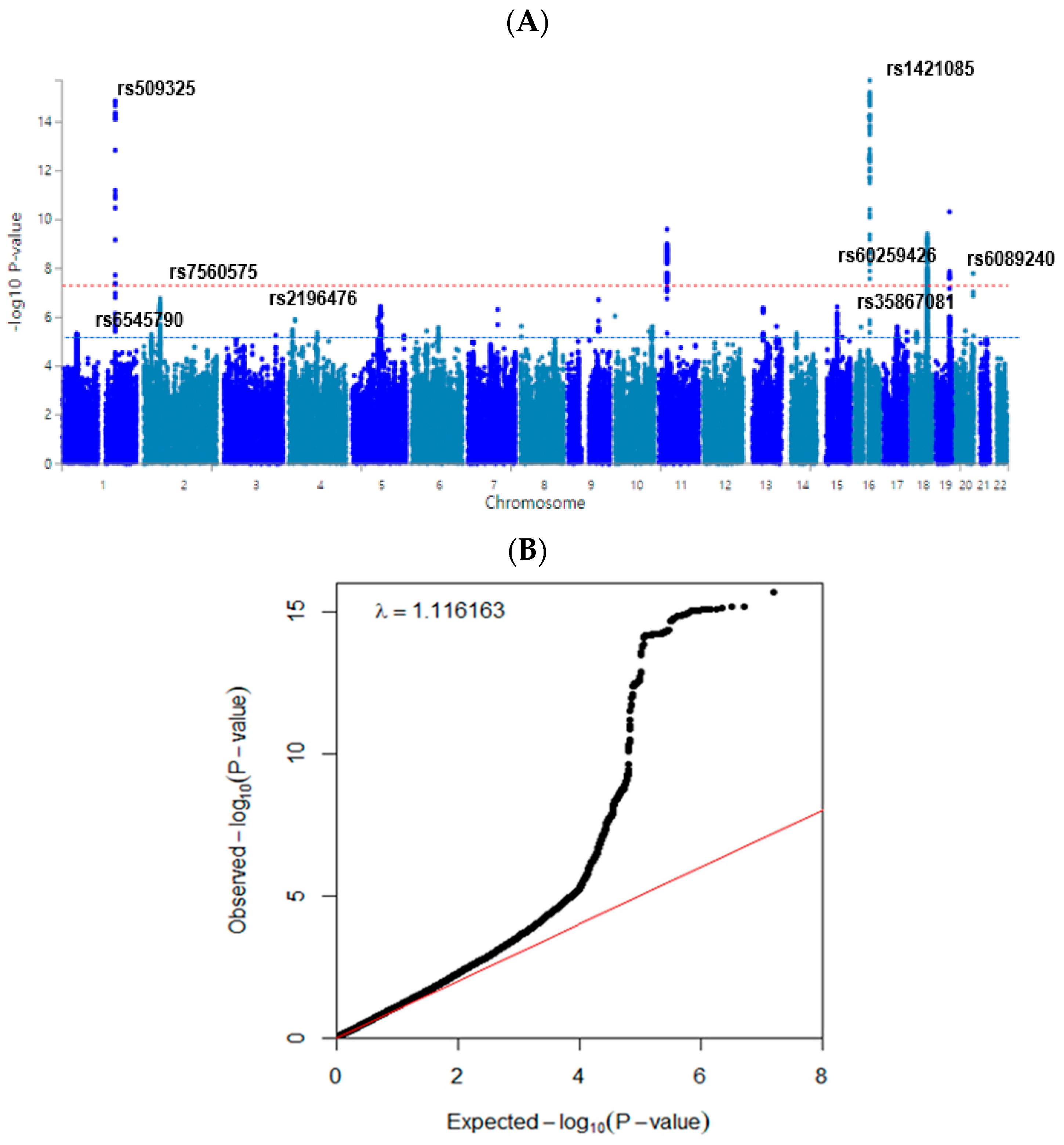
3.3. Polygenetic Variants and Their Interactions Related to Obesity Defined by Fat Mass
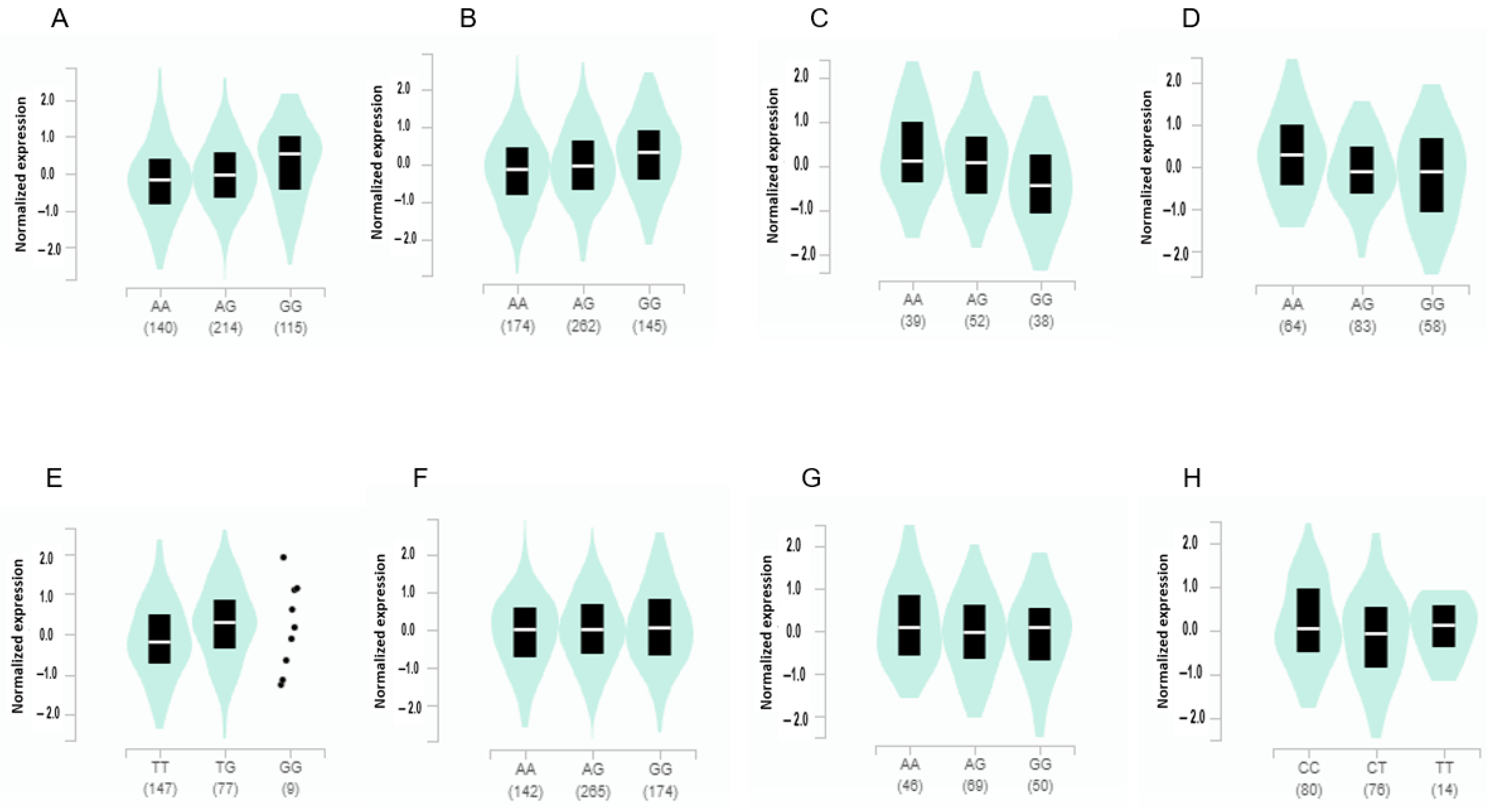
3.4. The Gene Expression According to the Alleles of the Selected SNPs in Different Tissues from GTEx v8
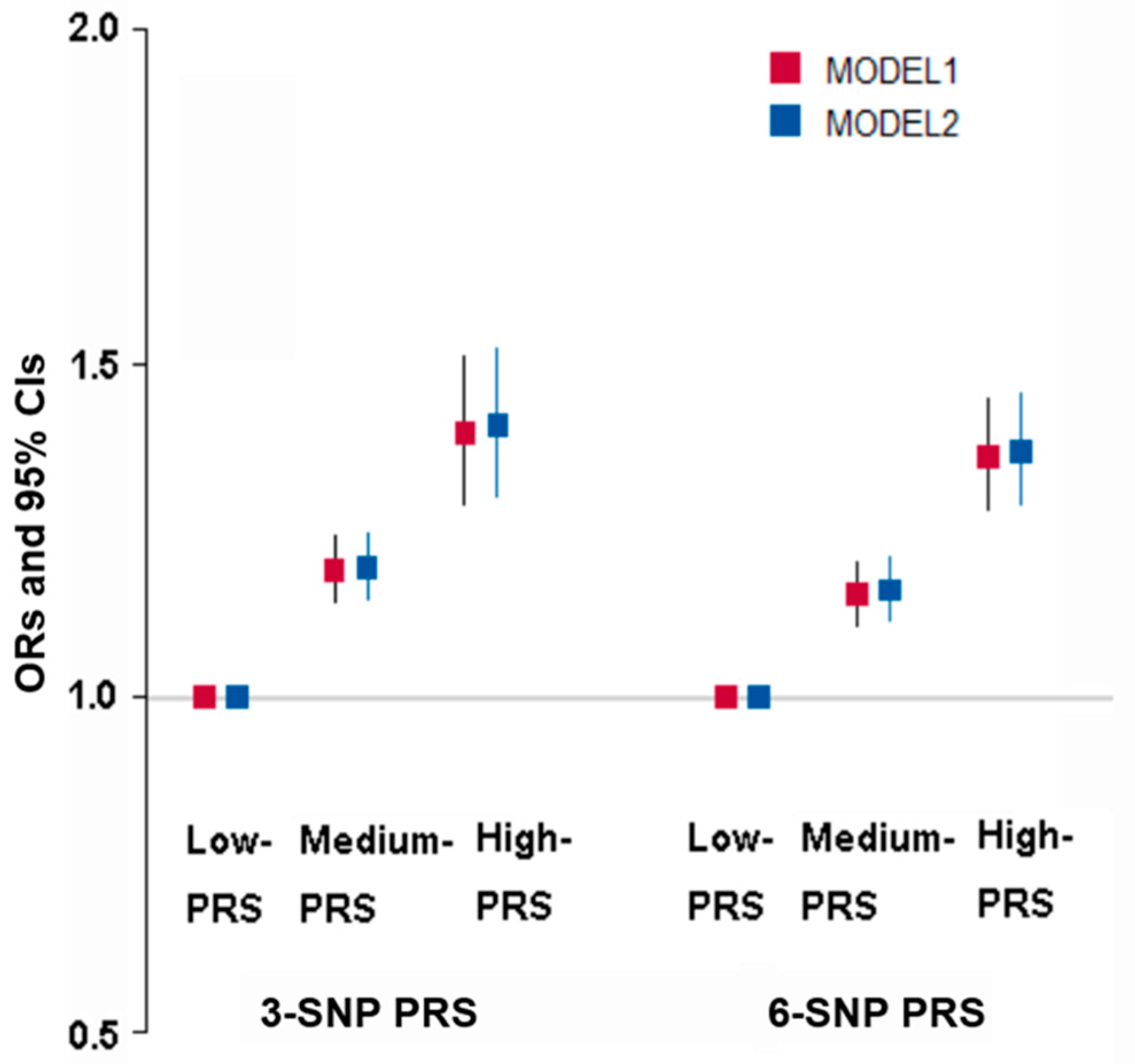
3.5. The Best Model of Genetic Variants with SNP–SNP Interaction for Obesity
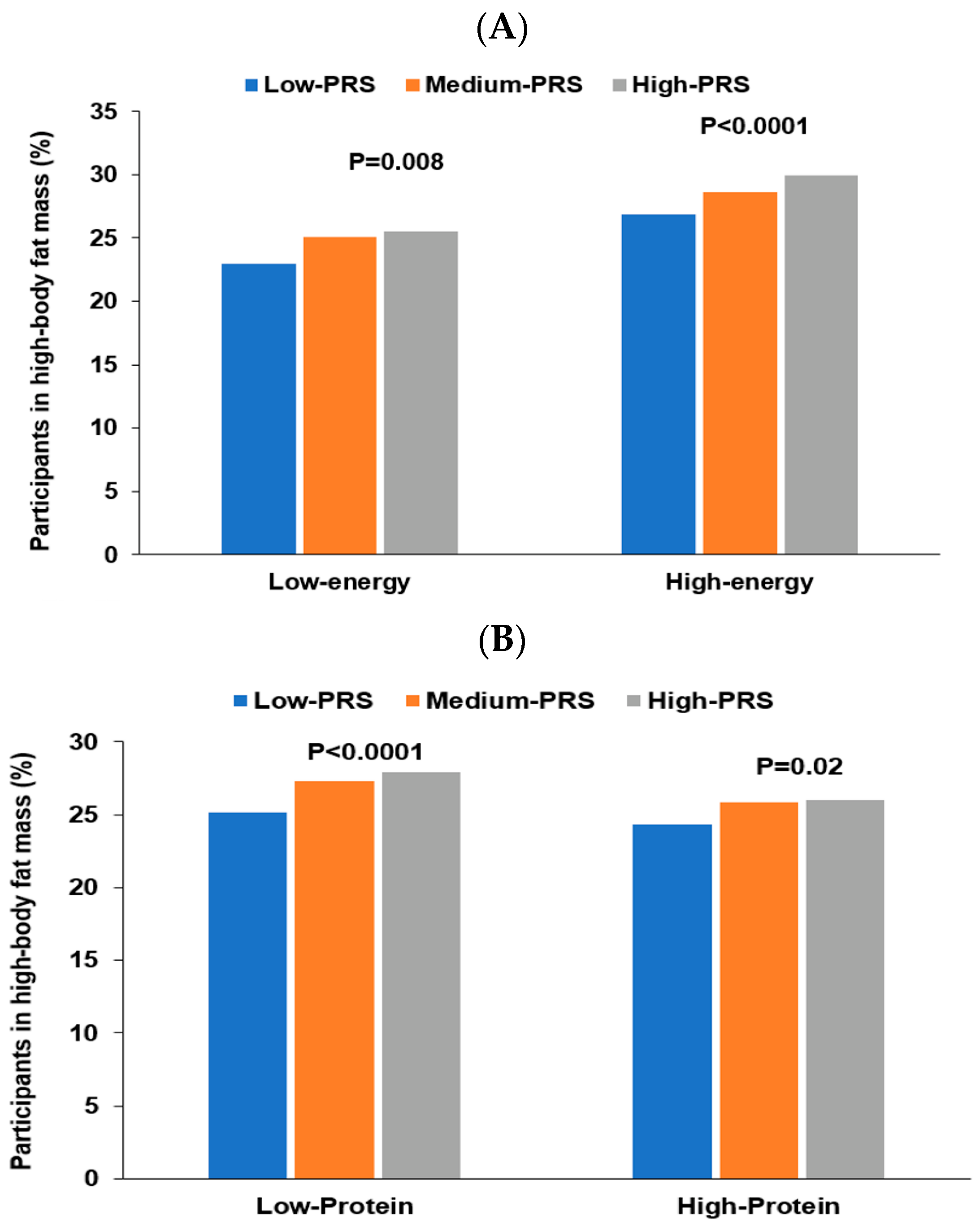
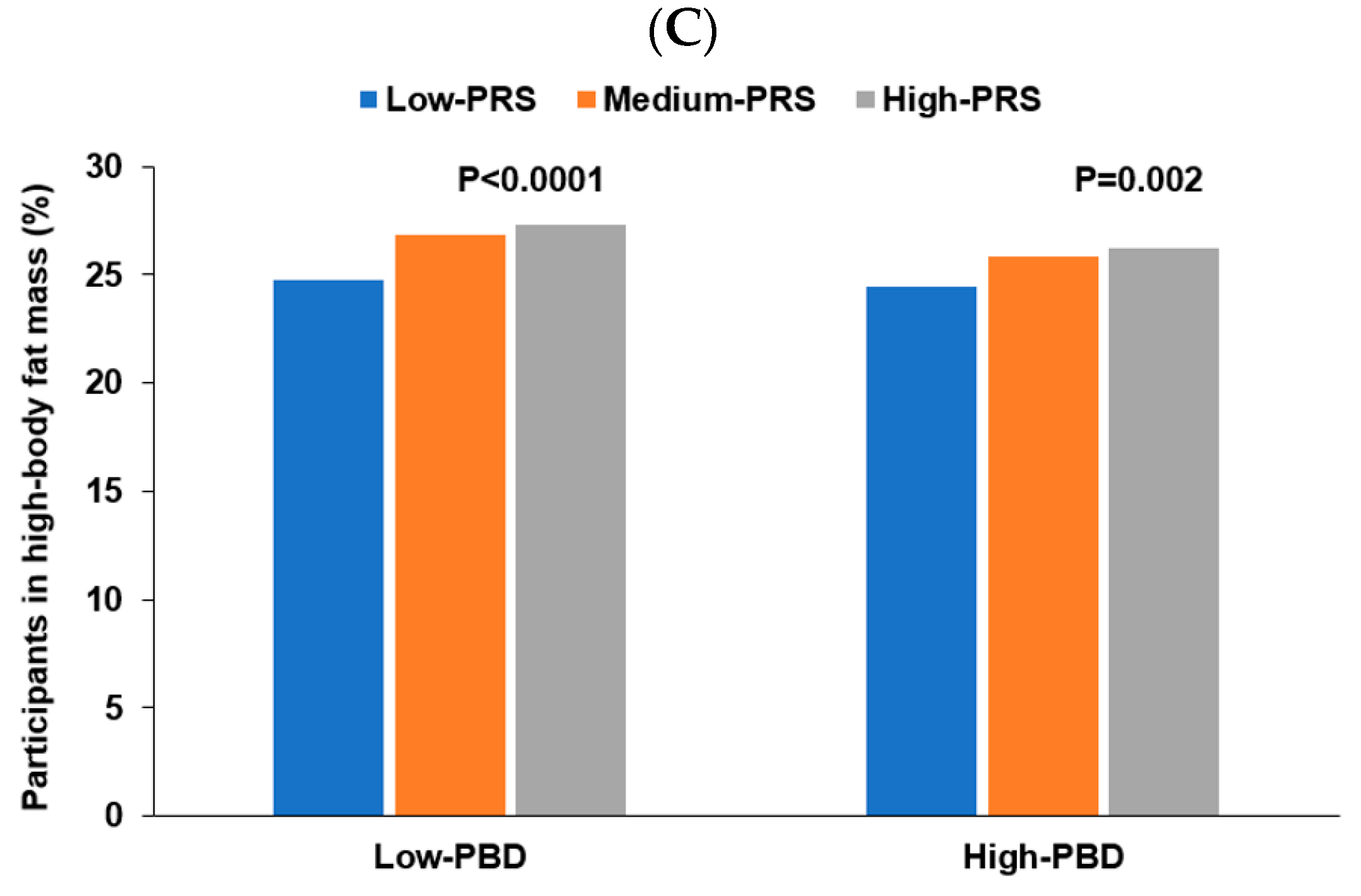
3.6. Interaction of PRS with Lifestyles to Influence Obesity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jacob, R.; Bertrand, C.; Llewellyn, C.; Couture, C.; Labonté, M.; Tremblay, A.; Bouchard, C.; Drapeau, V.; Pérusse, L. Dietary Mediators of the Genetic Susceptibility to Obesity-Results from the Quebec Family Study. J. Nutr. 2022, 152, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Yengo, L.; Sidorenko, J.; Kemper, K.E.; Zheng, Z.; Wood, A.R.; Weedon, M.N.; Frayling, T.M.; Hirschhorn, J.; Yang, J.; Visscher, P.M. Meta-analysis of genome-wide association studies for height and body mass index in ~700,000 individuals of European ancestry. Hum. Mol. Genet. 2018, 27, 3641–3649. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Yang, H.J.; Kim, M.J.; Hur, H.J.; Kim, S.H.; Kim, M.S. Interactions between Polygenic Risk Scores, Dietary Pattern, and Menarche Age with the Obesity Risk in a Large Hospital-Based Cohort. Nutrients 2021, 13, 3772. [Google Scholar] [CrossRef] [PubMed]
- Konuma, T.; Okada, Y. Statistical genetics and polygenic risk score for precision medicine. Inflamm. Regen. 2021, 41, 18. [Google Scholar] [CrossRef]
- Hu, F.B. 437Genetic Predictors of Obesity. In Obesity Epidemiology; Hu, F.B., Ed.; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Lim, H.; Kim, S.Y.; Wang, Y.; Lee, S.J.; Oh, K.; Sohn, C.Y.; Moon, Y.M.; Jee, S.H. Preservation of a traditional Korean dietary pattern and emergence of a fruit and dairy dietary pattern among adults in South Korea: Secular transitions in dietary patterns of a prospective study from 1998 to 2010. Nutr. Res. 2014, 34, 760–770. [Google Scholar] [CrossRef]
- Heymsfield, S.B.; Peterson, C.M.; Thomas, D.M.; Heo, M.; Schuna, J.M., Jr. Why are there race/ethnic differences in adult body mass index-adiposity relationships? A quantitative critical review. Obes. Rev. 2016, 17, 262–275. [Google Scholar] [CrossRef] [Green Version]
- Hamilton-James, K.; Collet, T.-H.; Pichard, C.; Genton, L.; Dupertuis, Y.M. Precision and accuracy of bioelectrical impedance analysis devices in supine versus standing position with or without retractable handle in Caucasian subjects. Clin. Nutr. ESPEN 2021, 45, 267–274. [Google Scholar] [CrossRef]
- Fang, W.-H.; Yang, J.-R.; Lin, C.-Y.; Hsiao, P.-J.; Tu, M.-Y.; Chen, C.-F.; Tsai, D.-J.; Su, W.; Huang, G.-S.; Chang, H.; et al. Accuracy augmentation of body composition measurement by bioelectrical impedance analyzer in elderly population. Medicine 2020, 99, e19103. [Google Scholar] [CrossRef]
- Kim, Y.; Han, B.G. Cohort Profile: The Korean Genome and Epidemiology Study (KoGES) Consortium. Int. J. Epidemiol. 2017, 46, e20. [Google Scholar] [CrossRef]
- Park, S. Association between polygenetic risk scores related to sarcopenia risk and their interactions with regular exercise in a large cohort of Korean adults. Clin. Nutr. 2021, 40, 5355–5364. [Google Scholar] [CrossRef]
- Wu, X.; Park, S. An Inverse Relation between Hyperglycemia and Skeletal Muscle Mass Predicted by Using a Machine Learning Approach in Middle-Aged and Older Adults in Large Cohorts. J. Clin. Med. 2021, 10, 2133. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, K.; Lee, B.K.; Ahn, J. A Healthy Diet Rich in Calcium and Vitamin C Is Inversely Associated with Metabolic Syndrome Risk in Korean Adults from the KNHANES 2013–2017. Nutrients 2021, 13, 1312. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.J.; Song, M.Y.; Kim, M.J.; Park, S. Associations between metabolic syndrome and urinary Na-to-K ratio and glomerular filtration rate in middle-aged adults regardless of Na and K intakes. Clin. Exp. Nephrol. 2020, 24, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Daily, J.W.; Song, M.Y.; Kwon, H.K. Gene-gene and gene-lifestyle interactions of AKAP11, KCNMA1, PUM1, SPTBN1, and EPDR1 on osteoporosis risk in middle-aged adults. Nutrition 2020, 79–80, 110859. [Google Scholar] [CrossRef]
- Park, S.; Kang, S. A Western-style diet interacts with genetic variants of the LDL receptor to hyper-LDL cholesterolemia in Korean adults. Public Health Nutr. 2021, 24, 2964–2974. [Google Scholar] [CrossRef]
- Ahn, Y.; Kwon, E.; Shim, J.E.; Park, M.K.; Joo, Y.; Kimm, K.; Park, C.; Kim, D.H. Validation and reproducibility of food frequency questionnaire for Korean genome epidemiologic study. Eur. J. Clin. Nutr. 2007, 61, 1435–1441. [Google Scholar] [CrossRef]
- Shivappa, N.; Steck, S.E.; Hurley, T.G.; Hussey, J.R.; Hébert, J.R. Designing and developing a literature-derived, population-based dietary inflammatory index. Public Health Nutr. 2014, 17, 1689–1696. [Google Scholar] [CrossRef] [Green Version]
- Hébert, J.R.; Shivappa, N.; Wirth, M.D.; Hussey, J.R.; Hurley, T.G. Perspective: The Dietary Inflammatory Index (DII)-Lessons Learned, Improvements Made, and Future Directions. Adv. Nutr. 2019, 10, 185–195. [Google Scholar] [CrossRef]
- Liu, M.; Park, S. A Causal Relationship between Vitamin C Intake with Hyperglycemia and Metabolic Syndrome Risk: A Two-Sample Mendelian Randomization Study. Antioxidants 2022, 11, 857. [Google Scholar] [CrossRef]
- Burns, K.F.; Robson, S.M.; Raynor, H.A. A low-energy-dense dietary pattern: A narrative review. Nutr. Rev. 2022, 80, 2301–2311. [Google Scholar] [CrossRef]
- Li, X.; Zheng, L.; Zhang, B.; Deng, Z.Y.; Luo, T. The Structure Basis of Phytochemicals as Metabolic Signals for Combating Obesity. Front. Nutr. 2022, 9, 913883. [Google Scholar] [CrossRef] [PubMed]
- Mou, Z.; Hyde, T.M.; Lipska, B.K.; Martinowich, K.; Wei, P.; Ong, C.J.; Hunter, L.A.; Palaguachi, G.I.; Morgun, E.; Teng, R.; et al. Human Obesity Associated with an Intronic SNP in the Brain-Derived Neurotrophic Factor Locus. Cell Rep. 2015, 13, 1073–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, V.T.; Hill, B.; Sims, N.; Heck, A.; Negron, M.; Lusk, C.; Galindo, C.L. Brain-derived neurotrophic factor rs6265 (Val66Met) single nucleotide polymorphism as a master modifier of human pathophysiology. Neural Regen. Res. 2023, 18, 102–106. [Google Scholar] [PubMed]
- Mozaffarian, D. Perspective: Obesity—And unexplained epidemic. Am. J. Clin. Nutr. 2022, 115, 1445–1450. [Google Scholar] [CrossRef] [PubMed]
- Daily, J.W.; Park, S. Interaction of BDNF rs6265 variants and energy and protein intake in the risk for glucose intolerance and type 2 diabetes in middle-aged adults. Nutrition 2017, 33, 187–194. [Google Scholar] [CrossRef]
- Ma, X.-Y.; Qiu, W.Q.; Smith, C.E.; Parnell, L.D.; Jiang, Z.-Y.; Ordovas, J.M.; Tucker, K.L.; Lai, C.-Q. Association between BDNF rs6265 and Obesity in the Boston Puerto Rican Health Study. J. Obes. 2012, 2012, 102942. [Google Scholar] [CrossRef] [Green Version]
- Smemo, S.; Tena, J.J.; Kim, K.H.; Gamazon, E.R.; Sakabe, N.J.; Gómez-Marín, C.; Aneas, I.; Credidio, F.L.; Sobreira, D.R.; Wasserman, N.F.; et al. Obesity-associated variants within FTO form long-range functional connections with IRX3. Nature 2014, 507, 371–375. [Google Scholar] [CrossRef] [Green Version]
- Claussnitzer, M.; Dankel, S.N.; Kim, K.-H.; Quon, G.; Meuleman, W.; Haugen, C.; Glunk, V.; Sousa, I.S.; Beaudry, J.L.; Puviindran, V.; et al. FTO Obesity Variant Circuitry and Adipocyte Browning in Humans. N. Engl. J. Med. 2015, 373, 895–907. [Google Scholar] [CrossRef] [Green Version]
- Laber, S.; Forcisi, S.; Bentley, L.; Petzold, J.; Moritz, F.; Smirnov, K.S.; Al Sadat, L.; Williamson, I.; Strobel, S.; Agnew, T.; et al. Linking the FTO obesity rs1421085 variant circuitry to cellular, metabolic, and organismal phenotypes in vivo. Sci. Adv. 2021, 7, eabg0108. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Murata, T. Involvement of regucalcin gene promoter region-related protein-p117, a transcription factor, in human obesity. Biomed. Rep. 2017, 6, 374–378. [Google Scholar] [CrossRef] [Green Version]
- Ng, M.C.; Tam, C.H.; So, W.Y.; Ho, J.S.; Chan, A.W.; Lee, H.M.; Wang, Y.; Lam, V.K.; Chan, J.C.; Ma, R.C. Implication of genetic variants near NEGR1, SEC16B, TMEM18, ETV5/DGKG, GNPDA2, LIN7C/BDNF, MTCH2, BCDIN3D/FAIM2, SH2B1, FTO, MC4R, and KCTD15 with obesity and type 2 diabetes in 7705 Chinese. J. Clin. Endocrinol. Metab. 2010, 95, 2418–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, D.; Glick, B.S. Two mammalian Sec16 homologues have nonredundant functions in endoplasmic reticulum (ER) export and transitional ER organization. Mol. Biol. Cell 2007, 18, 839–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domić, J.; Grootswagers, P.; van Loon, L.J.C.; de Groot, L. Perspective: Vegan Diets for Older Adults? A Perspective On the Potential Impact On Muscle Mass and Strength. Adv. Nutr. 2022, 13, 712–725. [Google Scholar] [CrossRef]
- Vang, A.; Singh, P.N.; Lee, J.W.; Haddad, E.H.; Brinegar, C.H. Meats, processed meats, obesity, weight gain and occurrence of diabetes among adults: Findings from Adventist Health Studies. Ann. Nutr. Metab. 2008, 52, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Fuente González, C.E.; Chávez-Servín, J.L.; de la Torre-Carbot, K.; Ronquillo González, D.; Aguilera Barreiro, M.; Ojeda Navarro, L.R. Relationship between Emotional Eating, Consumption of Hyperpalatable Energy-Dense Foods, and Indicators of Nutritional Status: A Systematic Review. J. Obes. 2022, 2022, 4243868. [Google Scholar] [CrossRef] [PubMed]
- Riedl, R.A.; Burnett, C.M.L.; Pearson, N.A.; Reho, J.J.; Mokadem, M.; Edwards, R.A.; Kindel, T.L.; Kirby, J.R.; Grobe, J.L. Gut Microbiota Represent a Major Thermogenic Biomass. Function 2021, 2, zqab019. [Google Scholar] [CrossRef]
- Armstrong, L.E.; Casa, D.J.; Belval, L.N. Metabolism, bioenergetics and thermal physiology: Influences of the human intestinal microbiota. Nutr. Res. Rev. 2019, 32, 205–217. [Google Scholar] [CrossRef]
- Li, D.; Li, Y.; Yang, S.; Lu, J.; Jin, X.; Wu, M. Diet-gut microbiota-epigenetics in metabolic diseases: From mechanisms to therapeutics. Biomed. Pharmacother. 2022, 153, 113290. [Google Scholar] [CrossRef]
- Mohamadi, A.; Shiraseb, F.; Mirzababaei, A.; Hosseininasab, D.; Rasaei, N.; Clark, C.C.T.; Mirzaei, K. Circulating Inflammatory Markers May Mediate the Relationship between Healthy Plant-Based Diet and Metabolic Phenotype Obesity in Women: A Cross-Sectional Study. Int. J. Clin. Pract. 2022, 2022, 8099382. [Google Scholar] [CrossRef]
- Im, J.; Kim, M.; Park, K. Association between the Phytochemical Index and Lower Prevalence of Obesity/Abdominal Obesity in Korean Adults. Nutrients 2020, 12, 2312. [Google Scholar] [CrossRef]
- Hansen, T.T.; Astrup, A.; Sjödin, A. Are Dietary Proteins the Key to Successful Body Weight Management? A Systematic Review and Meta-Analysis of Studies Assessing Body Weight Outcomes after Interventions with Increased Dietary Protein. Nutrients 2021, 13, 3193. [Google Scholar] [CrossRef] [PubMed]
- Binou, P.; Yanni, A.E.; Kartsioti, K.; Barmpagianni, A.; Konstantopoulos, P.; Karathanos, V.T.; Kokkinos, A. Wheat Biscuits Enriched with Plant-Based Protein Contribute to Weight Loss and Beneficial Metabolic Effects in Subjects with Overweight/Obesity. Nutrients 2022, 14, 2516. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Glenn, A.J.; Yang, Q.; Ding, D.; Zheng, L.; Bao, W.; Beasley, J.; LeBlanc, E.; Lo, K.; Manson, J.E.; et al. Dietary Protein Sources, Mediating Biomarkers, and Incidence of Type 2 Diabetes: Findings from the Women’s Health Initiative and the UK Biobank. Diabetes Care 2022, 45, 1742–1753. [Google Scholar] [CrossRef]
- Choi, M.K.; Bae, Y.J. Protein intake and osteosarcopenic adiposity in Korean adults aged 50 years and older. Osteoporos. Int. 2020, 31, 2363–2372. [Google Scholar] [CrossRef] [PubMed]
- Orlich, M.J.; Fraser, G.E. Vegetarian diets in the Adventist Health Study 2: A review of initial published findings. Am. J. Clin. Nutr. 2014, 100 (Suppl. S1), 353s–358s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slywitch, E.; Savalli, C.; Duarte, A.C.; Escrivão, M. Obese Vegetarians and Omnivores Show Different Metabolic Changes: Analysis of 1340 Individuals. Nutrients 2022, 14, 2204. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) | ||||||||
| Korean Balanced Diet | Plant-Based Diet | Western-Style Diet | Rice-Main Diet | |||||
| Rice | −3 | −7 | 6 | 93 | * | |||
| Whole grain | 8 | −4 | −3 | −93 | * | |||
| Bread | −1 | 5 | 63 | * | 1 | |||
| Cookie | −8 | 37 | 50 | −5 | ||||
| Noodles | −8 | 33 | 29 | * | 6 | |||
| Bean | 32 | 47 | * | 3 | 2 | |||
| Potato | 26 | 49 | * | 4 | −3 | |||
| Kimchi | 50 | * | 0 | 0 | −2 | |||
| Egg | 8 | 44 | * | 15 | 4 | |||
| Fast food | −5 | 18 | 73 | * | −4 | |||
| Green vegetables | 68 | * | 40 | * | −2 | −1 | ||
| White vegetables | 71 | * | 27 | 3 | 2 | |||
| Mushroom | 50 | * | 34 | −6 | −3 | |||
| Fatty fish | 53 | * | 21 | 12 | 0 | |||
| Whitefish | 66 | * | 15 | 15 | 0 | |||
| Crab | 48 | * | 3 | 21 | 1 | |||
| Processed meats | 18 | 15 | 7 | −1 | ||||
| Red meat | 43 | * | −6 | 44 | * | 7 | ||
| Chicken | 16 | 4 | 63 | −5 | ||||
| Soups | 32 | −5 | 40 | * | 3 | |||
| Seaweeds | 45 | * | 39 | * | −2 | −4 | ||
| Milk | 12 | 49 | * | 1 | 0 | |||
| Beverage | 20 | 31 | 6 | 2 | ||||
| Coffee | 9 | 0 | 20 | 14 | ||||
| Tea | 12 | −7 | 26 | 13 | ||||
| Fruits | 21 | 46 | * | −6 | −5 | |||
| Pickles | 50 | * | −2 | 6 | 2 | |||
| Alcohol | 16 | −27 | 18 | 5 | ||||
| Nuts | −1 | 50 | * | 5 | −5 | |||
| Variance explained by each dietary pattern | 3.57 | 2.44 | 2.30 | 1.79 | ||||
| (B) | ||||||||
| KBD | PBD | WSD | RMD | |||||
| Energy intake (EER%) | 125.6 ± 0.434 b | 109.8 ± 0.254 c | 178.9 ± 1.193 a | 87.1 ± 0.143 c | ||||
| Carbohydrates (En%) | 64.8_0.103 c | 69.4 ± 0.061 b | 63.7 ± 0.287 d | 73.2 ± 0.04 a | ||||
| Protein (EN%) | 17.1 ± 0.037 a | 14.0 ± 0.02 b | 14.8 ± 0.103 b | 12.8 ± 0.012 c | ||||
| Fat (En%) | 18.0 ± 0.08 b | 16.3 ± 0.047 c | 20.8 ± 0.221 a | 12.6 ± 0.02 d | ||||
| SFA (En%) | 4.15 + 0.13 d | 5.67 + 0.42 b | 4.89 + 0.24 c | 0.64 + 0.12 a | ||||
| MUFA (En%) | 5.18 + 0.15 d | 6.89 + 0.45 b | 6.13 + 0.257 c | 8.87 + 0.12 a | ||||
| PUFA (En%) | 2.93 + 0.16 d | 4.06 + 0.38 b | 3.51 + 0.22 c | 4.75 + 0.11 a | ||||
| Fiber (g) | 26.0 ± 0.123 | 15.1 ± 0.07 | 14.1 ± 0.34 | 13.3 ± 0.04 | ||||
| Vitamin C (mg) | 169.7 ± 0.885 a | 125.5 ± 0507 b | 54.5 ± 2.44 d | 91.6 ± 0.289 c | ||||
| Men (n = 19,444) | Women (n = 34,384) | |||
|---|---|---|---|---|
| Low-BF (n = 16,146) | High-BF (n = 3298) | Low-BF (n = 27,180) | High-BF (n = 7204) | |
| Age (year) | 55.7 ± 0.06 b | 56.9 ± 0.13 a | 51.9 ± 0.05 d | 53.8 ± 0.09 c***+++### |
| Education ≤ Middle school | 1318 (13.2) | 343 (16.8) ‡‡‡ | 4290 (20.3) | 1727 (28.4) ‡‡‡ |
| High school | 7540 (76.2) | 1491 (72.8) | 15,479 (73.4) | 4074 (67.0) |
| ≥College | 1040 (10.5) | 214 (10.5) | 1322 (6.27) | 276 (4.54) |
| Income | ||||
| ≤USD 2000 | 1258 (8.24) | 251 (7.76) ‡ | 2848 (11.2) | 868 (12.4) ‡‡‡ |
| USD 2000–4000 | 6565 (42.8) | 1311 (40.5) | 10,903 (43.0) | 3369 (48.0) |
| >USD 4000 | 7479 (49.0) | 1672 (51.7) | 11,579 (45.7) | 2777 (39.6) |
| Energy (EER%) | 86.0 ± 0.06 b | 85.4 ± 0.13 c | 104 ± 0.05 a | 104 ± 0.09 a***+++ |
| Carbohydrates (En%) | 71.6 ± 0.08 | 71.3 ± 0.17 | 71.7 ± 0.06 | 71.6 ± 0.12 ++ |
| Proteins (En%) | 13.3 ± 0.03 b | 13.4 ± 0.05 b | 13.6 ± 0.02 a | 13.6 ± 0.04 a***# |
| Fat (En%) | 13.9 ± 0.06 | 14.2 ± 0.12 | 13.9 ± 0.04 | 14.1 ± 0.09 |
| SFA (En%) | 4.46 ± 0.02 | 4.55 ± 0.06 | 4.45 ± 0.02 | 4.43 + 0.04 |
| MUFA (En%) | 5.62 ± 0.03 a | 5.80 ± 0.06 a | 5.45 ± 0.02 b | 5.48 ± 0.05 b***+ |
| PUFA (En%) | 3.26 ± 0.03 a | 3.21 ± 0.05 a | 3.12 ± 0.02 b | 3.07 ± 0.04 b** |
| Cholesterol (mg/d) | 169 ± 1.07 | 173 ± 2.25 | 170 ± 0.81 | 171 ± 1.52 |
| Vitamin C (mg/d) | 93.8 ± 0.68 c | 91.6 ± 1.50 c | 121 ± 0.53 a | 115 ± 1.05 b***++ |
| Vitamin D (ug/d) | 5.68 ± 0.05 c | 5.39 ± 0.10 d | 6.94 ± 0.04 a | 6.57 ± 0.07 b***+++ |
| Fiber (g/d) | 14.6 ± 0.09 b | 14.1 ± 0.20 b | 15.3 ± 0.07 a | 14.4 ± 0.14 b***+++ |
| DII (scores) | −19.96 ± 0.02 b | −21.07 ± 0.04 b | −21.26 ± 0.01 a | −21.34 ± 0.03 a**+# |
| Ca (mg/d) | 417 ± 3.42 b | 414 ± 5.56 b | 491 ± 2.31 a | 484 ± 3.81 a*** |
| KBD (Yes, %) | 6169 (38.2) | 1324(40.2) ‡ | 7379 (31.1) | 3057 (28.7) ‡‡‡ |
| PBD (Yes, %) | 3399 (21.1) | 693 (21.0) | 9675 (40.7) | 4136(38.9) ‡‡ |
| WSD (Yes, %) | 8218 (50.9) | 1877 (50.9) ‡‡‡ | 7871 (33.2) | 3684 (34.6) ‡‡ |
| RMD (Yes, %) | 5089 (31.5) | 1016 (30.8) | 8180 (34.5) | 3628 (34.1) |
| Non-Smokers (Yes, %) | 4712 (29.3) | 834 (25.3) ‡‡‡ | 22,864 (96.8) | 10,296 (96.9) |
| Former smokers (Yes, %) | 6770 (42.1) | 1546 (46.9) | 284 (1.2) | 132(1.24) |
| Smokers (Yes, %) | 4612 (28.7) | 917 (27.8) | 532 (1.97) | 203 (1.91) |
| Alcohol drinking (g/day) | 35.1 ± 0.40 b | 38.9 ± 0.84 a | 5.33 ± 0.29 c | 5.51 ± 0.56 c***+++### |
| Regular exercise (Yes, %) | 10155 (60.5) | 1797 (52.4) ‡‡‡ | 16350 (54.0) | 3674 (45.9) ‡‡‡ |
| Men (n = 19,444) | Women (n = 34,384) | Adjusted ORs and 95% CI | |||
|---|---|---|---|---|---|
| Low-BF (n = 16,146) | High-BF (n = 3298) | Low-BF (n = 27,180) | High-BF (n = 7204) | ||
| Height (cm) 1 | 169.0 ± 0.04 b | 167.1 ± 0.09 a | 157.0 ± 0.03 d | 154.9 ± 0.07 c***+++# | 3.093 (2.881–3.320) |
| BMI (mg/kg2) 2 | 23.9 ± 0.02 c | 26.9 ± 0.05 a | 22.9 ± 0.02 d | 26.1 ± 0.03 b***###+++ | 18.05 (17.03–19.14) |
| Waist (cm) 3 | 84.3 ± 0.06 c | 91.7 ± 0.13 a | 76.7 ± 0.05 d | 83.4 ± 0.09 b***###+++ | 5.038 (4.794 5.293) |
| SMI (%) 4 | 7.84 ± 0.01 a | 7.50 ± 0.01 b | 7.05 ± 0.003 c | 6.68 ± 0.006 d***###+++ | 0.858 (0.745–0.954) |
| Fat mass (%) 5 | 22.4 ± 0.05 d | 26.3 ± 0.09 c | 29.8 ± 0.04 b | 33.9 ± 0.06 a***+++ | |
| MetS (%) 6 | 2393 (14.2) | 1205 (35.1) ‡‡‡ | 2933 (9.65) | 1769 (22.1) ‡‡‡ | 6.289 (5.833–6.780) |
| glucose (mg/dL) 7 | 98.2 ± 0.17 b | 99.6 ± 0.37 a | 93.2 ± 0.13 d | 94.1 ± 0.25 c***+++ | 1.178 (1.103–1.257) |
| HbA1c (%) 8 | 5.72 ± 0.01 b | 5.77 ± 0.01 a | 5.70 ± 0.01 b | 5.69 ± 0.01 b***## | 1.454 (1.321–1.601) |
| Total cholesterol 9 | 190.4 ± 0.31 d | 194.9 ± 0.68 c | 199.4 ± 0.24 b | 207.3 ± 0.46 a***++ | 1.073 (1.001–1.150) |
| HDL (mg/dL) 10 | 49.2 ± 0.11 c | 49.8 ± 0.24 c | 55.8 ± 0.08 a | 57.6 ± 0.16 b***+++### | 1.210 (1.135–1.290) |
| LDL (mg/dL) 11 | 113 ± 0.28 b | 114 ± 0.62 c | 121 ± 0.22 a | 126 ± 0.42 a***+++### | 1.089 (1.007–1.177) |
| TG (mg/dL) 12 | 142 ± 0.72 b | 158 ± 1.59 a | 115 ± 0.57 c | 117 ± 1.07 c***+++### | 1.229 (1.156–1.308) |
| Hs-CRP (mg/dL) 13 | 0.153 ± 0.004 b | 0.187 ± 0.007 a | 0.118 ± 0.003 c | 0.154 ± 0.005 b***+++ | 1.266 (1.008–1.589) |
| SBP (mmHg) 14 | 125.0 ± 0.14 a | 124.9 ± 0.24 a | 121.0 ± 0.10 b | 121.4 ± 0.20 b*** | 1.105 (1.040–1.175) |
| DBP (mmHg) 15 | 78.1 ± 0.10 a | 77.9 ± 0.16 a | 74.4 ± 0.07 b | 74.6 ± 0.13 b***# | 1.040 (0.948–1.140) |
| eGFR (mL/min) 16 | 84.5 ± 0.16 b | 83.0 ± 0.26 c | 87.2 ± 0.11 a | 87.7 ± 0.23 a***+### | 0.774 (0.687–0.873) |
| AST (U/L) 17 | 24.6 ± 0.24 b | 25.7 ± 0.40 a | 23.1 ± 0.16 c | 23.2 ± 0.34 c**+++ | 1.425 (1.250–1.624) |
| ALT(U/L) 18 | 25.1 ± 0.24 b | 27.9 ± 0.39 a | 20.5 ± 0.16 c | 20.9 ± 0.33 b***+++### | 1.333 (1.227–1.448) |
| Pathways | No. of Genes | Beta | Std | p-Value | p-Value for Bonferroni Correction |
|---|---|---|---|---|---|
| GO BP: GO sodium ion transmembrane transport | 133 | 0.323 | 0.027 | 0.081 | 3.18 × 10−5 |
| GO BP: GO myoblast proliferation | 17 | 0.693 | 0.021 | 0.179 | 5.41 × 10−5 |
| Curated gene sets: Biocarta flumazenil pathway | 8 | 1.208 | 0.025 | 0.326 | 0.000104 |
| GO BP: GO Neuron apoptotic process | 224 | 0.211 | 0.023 | 0.058 | 0.000129 |
| Curated gene sets: Reactome RMTS methylate histone arginine | 60 | 0.387 | 0.022 | 0.108 | 0.000172 |
| GO MF: GO Peptide hormone binding | 47 | 0.488 | 0.024 | 0.137 | 0.000183 |
| GO BP: GO Positive regulation of vascular endothelial cell proliferation | 13 | 0.778 | 0.020 | 0.219 | 0.000193 |
| CHR 1 | SNP 2 | Location | Mi 3 | Ma 4 | OR 5 | SE 6 | p-Value for OR 7 | p-Value for OR 8 | Genes | Feature | 9 MAF | 10 HWE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | rs509325 | 177894591 | G | T | 1.123 | 0.0145 | 1.37 × 10−15 | 8.28 × 10−4 | SEC16B | Intron | 0.2845 | 0.5035 |
| 2 | rs6545790 | 25109302 | A | G | 1.063 | 0.0133 | 4.87 × 10−6 | 4.75 × 10−4 | ADCY3 | Intron | 0.4374 | 0.4064 |
| 2 | rs7560575 | 54142030 | C | T | 0.9889 | 0.0670 | 2.33 × 10−7 | 1.94 × 10−2 | PSME4 | Transcript | 0.0112 | 0.2631 |
| 4 | rs2196476 | 20270600 | G | A | 1.097 | 0.0191 | 1.26 × 10−6 | 0.0069 | SLIT2 | Intron | 0.135 | 0.4579 |
| 11 | rs6265 | 27679916 | C | T | 0.923 | 0.0133 | 2.45 × 10−10 | 0.0048 | BDNF | Missense | 0.4588 | 0.1481 |
| 13 | rs587056 | 98976374 | T | C | 1.35 | 0.0636 | 2.33 × 10−10 | 0.0067 | FARP1 | Intron | 0.0106 | 0.5496 |
| 16 | rs1421085 | 53800954 | T | C | 1.173 | 0.0197 | 6.24 × 10−16 | 2.82 × 10−6 | FTO | Transcript | 0.1245 | 0.4604 |
| 17 | rs35867081 | 79047278 | G | A | 1.065 | 0.0137 | 3.99 × 10−6 | 0.0052 | BAIAP2 | Transcript | 0.3651 | 0.2094 |
| 19 | rs60259426 | 46340832 | G | A | 0.938 | 0.0134 | 1.54 × 10−7 | 0.00072 | SYMPK | Transcript | 0.4202 | 0.3135 |
| 20 | rs6089240 | 60152260 | A | G | 0.9278 | 0.0126 | 1.60 × 10−8 | 5.81 × 10−4 | CDH4 | Intron | 0.4641 | 1.0 |
| Low-PRS (n = 19,686) | Medium-PRS (n = 30,513) | High-PRS (n = 3629) | Gene–Nutrient Interaction p-Value | |
|---|---|---|---|---|
| Low energy 1 High energy | 1 | 1.150 (1.081–1.223) 1.102 (1.021–1.189) | 1.228 (1.088–1.387) 1.157 (0.997–1.348) | 0.0231 |
| Low CHO 2 70 High CHO | 1 | 1.045 (0.893–1.223) 0.991 (0.927–1.060) | 1.006 (0.866–1.169) 1.079 (1.014–1.149) | 0.3493 |
| Low protein 3 13 High protein | 1 | 1.123 (1.052–1.198) 0.961 (0.881–1.049) | 1.255 (1.103–1.427) 1.019 (0.938–1.107) | 0.0272 |
| Low fat 4 15 Moderate fat | 1 | 1.000 (0.926–1.080) 0.994 (0.896–1.102) | 1.088 (1.013–1.169) 1.028 (0.932–1.133) | 0.0664 |
| Low alcohol 5 20 High alcohol | 1 | 1.139 (1.075–1.208) 1.129 (1.050–1.213) | 1.218 (1.086–1.366) 1.204 (1.046–1.386) | 0.8083 |
| Low KBD 6 High KBD | 1 | 0.997 (0.938–1.061) 1.002 (0.929–1.080) | 1.067 (1.007–1.130) 1.083 (1.009–1.162) | 0.5368 |
| Low PBD 6 High PBD | 1 | 1.132 (1.081–1.185) 1.023 (0.961–1.088) | 1.258 (1.126–1.406) 1.110 (1.006–1.224) | 0.0026 |
| Low WSD 6 High WSD | 1 | 0.997 (0.938–1.061) 0.998 (0.927–1.075) | 1.067 (1.007–1.130) 1.074 (1.002–1.151) | 0.1356 |
| Low RMD 6 High RMD | 1 | 1.112 (1.057–1.169) 1.159 (1.096–1.225) | 1.197 (1.134–1.263) 1.254 (1.128–1.395) | 0.8419 |
| Low exercise 7 High exercise | 1 | 1.034 (0.949–1.128) 0.997 (0.938–1.061) | 1.103 (1.018–1.196) 1.067 (1.007–1.130) | 0.1778 |
| Non-smoke 8 Smoke | 1 | 0.997 (0.938–1.061) 1.071 (0.948–1.210) | 1.067 (1.007–1.130) 1.041 (0.926–1.169) | 0.1328 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daily, J.W.; Park, S. Association of Plant-Based and High-Protein Diets with a Lower Obesity Risk Defined by Fat Mass in Middle-Aged and Elderly Persons with a High Genetic Risk of Obesity. Nutrients 2023, 15, 1063. https://doi.org/10.3390/nu15041063
Daily JW, Park S. Association of Plant-Based and High-Protein Diets with a Lower Obesity Risk Defined by Fat Mass in Middle-Aged and Elderly Persons with a High Genetic Risk of Obesity. Nutrients. 2023; 15(4):1063. https://doi.org/10.3390/nu15041063
Chicago/Turabian StyleDaily, James W., and Sunmin Park. 2023. "Association of Plant-Based and High-Protein Diets with a Lower Obesity Risk Defined by Fat Mass in Middle-Aged and Elderly Persons with a High Genetic Risk of Obesity" Nutrients 15, no. 4: 1063. https://doi.org/10.3390/nu15041063